free
trial issue
subscribe
back
issues
Athena
Review Vol. 5, no. 1
Records
of Life: Fossils as
Original
Sources
Permian
Russia
Early
Geologic mappings of western Russia
The geologic area from St.
Petersburg to the Ural Mountains (an area now known as the East
European Platform), flanked
on the north by
the Baltic
and Berents Sea coastline, was
first studied in detail in the early 19th
century. The initial geological map of the region, made by Strangeways
(1822),
provided an important starting point although it contained
innaccuracies. In the same period, the biologist Heinz
Christian Pander
(1830, 1856, 1857) made many
important discoveries of Baltic marine fossils from the Silurian and
Devonian periods.
Permian strata along the North Dvina River, which
flows north into the White Sea at Arkhangelsk, were
initially identified
by the Scottish geologist Roderick
Murchison in 1840-1. This came
in the process of defining the
geologic
interval between the Carboniferous and Triassic as the Permian period,
named
after the ancient kingdom of Permia
(Murchison et al. 1845).
 Murchison had previously
defined
the Silurian period from fossiliferous strata in England and Wales, and
then helped to identify the subsequent Devonian period (Murchison
1839). In the process, he was
able to show that various fossils in the Devonian strata of
Britain
were equivalent to those recently described in Russia by
Pander.
Murchison had previously
defined
the Silurian period from fossiliferous strata in England and Wales, and
then helped to identify the subsequent Devonian period (Murchison
1839). In the process, he was
able to show that various fossils in the Devonian strata of
Britain
were equivalent to those recently described in Russia by
Pander.
These included
the lobe-finned
fish Holoptychius nobilissimus,
first defined by
Louis Agassiz,
now
placed in the porolepiform order
established by
Swedish paleontologist Erik Jarvik. Although
closer to lungfishes (Dipnoi) than to the direct ancestors of
tetrapods, one
genus of porolepiforms named Laccognathus,
recently found
in
both Latvia and Ellesmere Island was
apparently
amphibious along rivers (see section on tetrapods for more details).
In 1840-1841 Murchison and
two colleagues,
E.
de
Verneuil and A. von Keyserling, and a Russian minerologist, Lieutenant
Koksharof, explored
the geological
formations east of St.
Petersburg and Moscow
(Murchison et al. 1845).
De Verneiul, a French expert on molluscs,
had previously worked with Murchison on definining the Devonian
sequence
in the Rhine Valley.
Fig.1:
Map of Permian and Mesozoic geologic formations west of the Ural
Mountains.
One of the main Permian zones
they
encountered lay within the flat topography of Northwestern
Russia,
along the Dvinia River (fig.1).
On ascending the
Dvina River from Arkhangelsk, Murchison and his colleagues recognized
Permian
formations in
riverbank exposures, sometimes comprising alternating layers of white
gypsum
and reddish or oxidized sandstone, representing riverine deposits
overlying marine limestone
strata.
At this time, both marine and freshwater bivalve fossils were recovered
by de Verneiul from
the
limestone layers (Murchison et al. 1845). In the 170 years since their
initial definition of the Permian period, the fossil record of the
Russian or East European block has been increasingly studied, and has
provided one of the world's most important zones of evidence
on paleozoic and mesozoic vertebrate evolution.

Permian
stratigraphy of the Russian
Platform
The Permian
in Russia is subdivided into three
stages: Ufimian, Kazanian, and Tatarian, which are further subdivided
into horizons, tetrapod zones, and faunal assemblages (fig.2).
The
type sections of the Upper Permian are
located in the eastern part of the Russian (East European) Platform and
the
Fore-Urals Trough in Tatarstan and Bashkortostan. In terms of overall
geology, the Russian
platform
is underlain by a crystalline basement composed of Archean
and Proterozoic rocks which are located
at depths of 1500 to 2000 m and more. This is overlain by a sedimentary
sheath
composed (successively) of Devonian, Carboniferous, Permian, Jurassic,
Cretaceous, Neogene and Quaternary rocks (Silantiev et al. 2013).
Fig.2:
Permian Stages and Horizons in East European Russia, and their
corresponding tetrapod zones and faunal assemblages (after Sennikov
and
Golubev
2005) .
Ufimian
stage of the Permian in Russia
The
Ufimian stage of the Permian is
subdivided into two horizens, the Solikamskian and the Shemshian. The
lowermost
is named for the town of Solikamsk, located near Perm, in the trough
along the
Urals.
Here the type section of this
horizon contains Lower and Upper subformations.
The Lower Solikamskaya Subformation,
170-200 m
thick, consists of
deeply buried dark-grey
clays and marl which have no outcrops, and must be studied in
boreholes. It
conformably overlies the Kungurian strata, and its start is marked by
the
appearance of the non-marine bivalves Palaeomutela
and Concinella,
both absent in the Lower Permian in this area. It also has a few thin
interbeds
with rare marine bivalves of the genera Schizodus,
Netschajewia,
and Permorphus
(Silantiev et al. 2013).
 The
mussel-shaped, freshwater bivalve
Palaeomutala was first
identified by
Amalitzky in 1892 on the basis of its distinctive multi-toothed hinges
(fig.3).
It is widespread throughout the Upper Permian strata of the Russian
Block. The
shells are oval or oval-rectangular, similar to those of modern
freshwater mussels
in the order Unionoida, which burrow into the substrate and eat
plankton and
other dissolved organic matter.
The
mussel-shaped, freshwater bivalve
Palaeomutala was first
identified by
Amalitzky in 1892 on the basis of its distinctive multi-toothed hinges
(fig.3).
It is widespread throughout the Upper Permian strata of the Russian
Block. The
shells are oval or oval-rectangular, similar to those of modern
freshwater mussels
in the order Unionoida, which burrow into the substrate and eat
plankton and
other dissolved organic matter.
Other
distinctive traits of the Palaeomutala
genus, such as a cross-lamellar
structure in the middle of its three shell layers, help secure its
identification,
making it a reliable indicator of freshwater bays and riverine settings
during
the Permian (Silantiev 1998). Based on multi-level sampling in
both
outcrops and wells, there are currently a number of proposed
species, each
correlated with different freshwater environments, ranging from water
with fast
currents to calm waters, and with fish and tetrapod fossil occurrences
(Silantiev 2014). Palaeomutala
is also found in Late Permian strata in
China.
Fig.3:
The mussel-like Palaeomutala,
commonly found in Permian fresh water environments in the Russian
Block (after Silantiev 1998). Its distinctive hinge mechanism with
numerous tooth-like elements is shown.
The
clam-shaped bivalve Concinella
(fig.4) is also commonly found in Permian strata
representing freshwater sites. Its type locations are along
the Pechora
River in Timan-Pechora
Province, and in the Kuznetsk basin (Pogorovich 1977).

The
Upper Solikamskaya Subformation, 50-240 m in
thickness, is exposed on the right bank of the Kama River near the
village of
Tyulkino, 25 km north of Solikamsk,.This subformation is composed of
alternating clays, marl, and limestones with abundant non-marine fauna
of
ostracodes and bivalves (Palaeomutela,
Sinomya,
Redikorella,
and Concinella),
and also with the remains of insects, fishes, and
plants (Silantiev 1996).
The
western edge of the Upper Solikamskaya
Subformation also outcrops along the valleys of the Vyatka and Sheshma
Rivers. Here
it is composed mainly of grey marl and dolomite, up to 10–15
m thick. Three of
the four non-marine bivalves already noted (Palaeomutela,
Redikorella,
and Concinella)
also occur here, although rarely.
Fig.4:
Casts of the clam-like freshwater bivalve Concinella,
whose fossils
are found
in numerous Ufimian deposits (after Silantiev 1998) .
The
later,
Sheshmian Horizon of the Ufimian
Stage outcrops in the lower part of the banks of the Kama
River
(upstream of
the mouth of the Vyatka River), and in the basin of the Shemsha River,
from which it received its name. This
horizon is largely composed
of red-bed sandstones, which are usually recognized as
alluvial
formations.
Fossils include
rare fish remains, leaf imprints, pollen and spores, and numerous
non-marine bivalves including Palaeomutela
and Concinella,
both
continuing
from the previous Solikamskian horizon; and ostracodes, the tiny
shrimp-like
crustaceans often used as relatively precise stratigraphic or horizon
markers. The
Sheshmian Horizon is also associated with the tetrapod Clamorosaurus
nocturus, the
type fauna for the Inta Faunal Assemblage (fig.2; cf.
Sennikov and
Golubev
2005).
Clamorosaurus
(fig.5)
was
a small temnospondyl amphibian about 23 cm (9
inches) long, dated at 273-272 mya. It
was a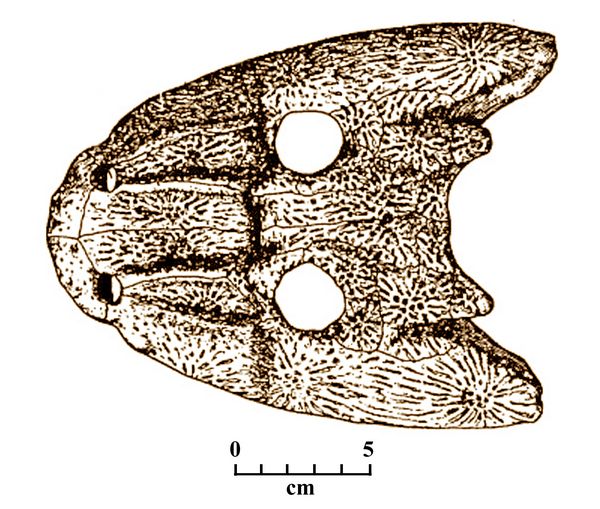 member of the
Eryopidae family, named for the type genus Eryops,
which
was first discovered in the Early Permian red beds of northern Texas
(Cope
1882). Besides
its very wide, shield-shaped skull,
Clamorosaurus
has large external nares or nostrils; both
features are common to all members of the Eryopidae family.
Comparisons
have often been made between Eryopidae fossils found from
the Ufimian stage of the Permian in the Russian Block, and
those in
the North
American Permian basins of Texas, Oklahoma, and New Mexico, where
Eryopidae
are well represented in the
latest stages of the Early Permian.
member of the
Eryopidae family, named for the type genus Eryops,
which
was first discovered in the Early Permian red beds of northern Texas
(Cope
1882). Besides
its very wide, shield-shaped skull,
Clamorosaurus
has large external nares or nostrils; both
features are common to all members of the Eryopidae family.
Comparisons
have often been made between Eryopidae fossils found from
the Ufimian stage of the Permian in the Russian Block, and
those in
the North
American Permian basins of Texas, Oklahoma, and New Mexico, where
Eryopidae
are well represented in the
latest stages of the Early Permian.
Fig.5:
Skull of Clamorosaurus seen
from above,
showing its wide profile and relatively large nostril openings, and the
dermal scales
which functioned as a protective surface on the skull.
.
Kazanian
stage of the Permian
The
second stage
of the Russian Permian, the Kazanian, begins with the lower Sokian
horizon (named for the Sok River) which mainly
outcrops in the eastern part of Tatarstan, along the Kama River, and in
the
basins of
the Sheshma, Zai, Menzela, and Ik rivers (Gusev, Burov, Esaulova et al.
1993).
The Sokian horizon represents a marine incursion which covered lower
elevations. It is composed of three formations of calcareous
clays, each 10-40 m thick, containing fossils
of
shallow marine
organisms including bivalves, bryozoans, brachiopods, crinoids,
ostracodes, and
foraminifers (Silantiev et al. 2013).
The
Upper Kazanian substage, widespread in Taterstan, is called the
Povolzhian
horizon. Its lowest strata, called
the Prikazansky formation, 45-50 m
thick, begins with a
limestone bed with a typical Lower Kazanian marine fauna including
brachiopods and crinoids.
This changes to terrestial conditions in the higher formations of the
Upper Kazanian stage, where a transition zone of clayish-marly
rocks contain mostly non-marine bivalves as well
as terrestrial plant remains. Marine and lagoonal facies of
the
Upper
Kazanian occur
on the right bank of the Volga River, near the villages of Pechishchi,
Naberezhye Morkvashi, and Krasnovidovo, while continental facies occur
on the
Kama River near the towns of Sheremetyevka and Sentyak.The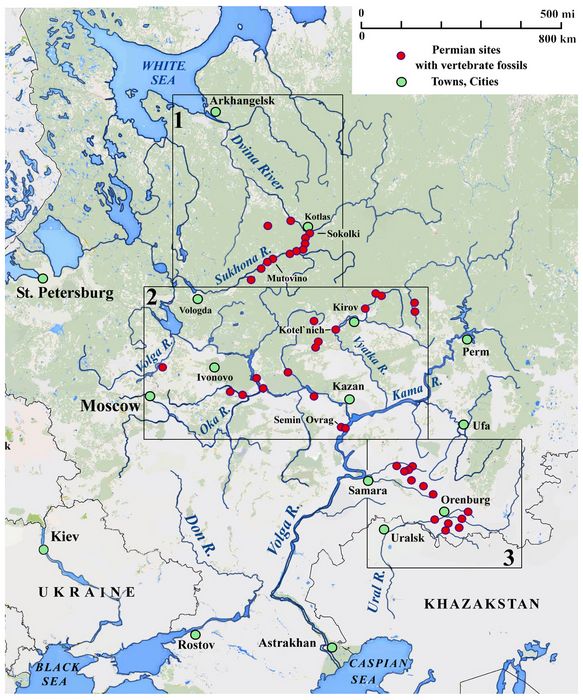 Belebeevskaya
Formation, some 60-100 m thick,
is composed of reddish-brown siltstones
and sandstones which contains a mixed fauna including the freshwater
mollusc Palaeomutela
umbonata,
fish
scales and teeth, fragments of reptile bones, imprints of leaves and
trees, and plant pollen and spores (Silantiev
et
al. 2013).
Belebeevskaya
Formation, some 60-100 m thick,
is composed of reddish-brown siltstones
and sandstones which contains a mixed fauna including the freshwater
mollusc Palaeomutela
umbonata,
fish
scales and teeth, fragments of reptile bones, imprints of leaves and
trees, and plant pollen and spores (Silantiev
et
al. 2013).
Tatarian
stage
of the Permian
The
third and highest stage of the Permian,
the Tatarian
stage, represents a terrestial setting which was crossed by numerous
rivers. This widespread
stage, up to 200-250 m in thickness, is mainly exposed in in the
watersheds of
rivers, almost always overlying the eroded surface of the Kazanian. The
Tatarian is
composed of red-bed (variegated) clays, siltstones, sandstones, marls,
dolomites, and limestones. These contain only non-marine fossils
including
bivalves, plants, insects, and vertebrates. The type sections are
located
outside Tatarstan, on the Vyatka River between the mouth of the Cheptsa
River
and the mouth of the Kobra River.
Fig.6:
Middle and Late Permian sites along three river drainages in western
Russia,
including 1) the North Dvina River; 2) the Volga River; and 3) the Ural
River (after
Golubev 2000a, fig.6.).
The
Tatarian is subdivided into Lower,
Middle,
and Upper substages,
corresponding with the Urzhuminian, Severodvinian,
and Vyatkian horizons. These have extensive fossil deposits of
terrestial flora and fauna, which will be explored in several site
areas
described below.
Today, about 170 years
after the initial exploration of Permian deposits in Russia by
Murchison et al. (1845),
dozens of large Tatarian sites haved been mapped and intensely
investigated along the
drainages of the North Dvina, Volga, and Ural Rivers (fig.6, boxes
1-3). The
Tatarian
vertebrate fossils found
west of the Urals have been grouped
into several faunal assemblages
(fig 2, and Golubev 2000a). The Middle-to-Late Tatarian phase, known as
the Severodvinian Horizon, is defined on the basis of faunal
assemblages from three highly productive sites
including Kotel'nich, Ilyinskoe (Semin Ovrag), and Sokolki,
together grouped as the Sokolki Assemblage.
This is followed chronologically
by the Vayazniki Assemblage,
representing the very end of the Tatarian up to the
Permian-Triassic extinction. Together these provide the
Russian equivalent of the Middle to Late Permian, Pristerognathus
through Dicynodon
Faunal
assemblages in South
Africa, and are
also comparable
to the Rio de Rasto faunal assemblage
in Brazil
(Cisneros et al. 2005).
.
Middle
Permian fossil deposits at Kotel'nich
 Along a
relatively straight, south-flowing stretch of the Vyatka River, about
80 km southwest of Kirov, significant Permian
layers with fossil deposits of an age corresponding to the Late
Capitanian stage are exposed near the town
of Kotel`nich. These Middle Permian red beds, composed
of
mudstones, siltstones, marl, sandstones,
and conglomerates, are exposed on the western bank of the Vyatka river
for 24
km between the towns of Kotel`nich and Zemtsy (fig.7). The
Vytaka River is a tributary of the Kama River, which, in turn, is one
of the main branches of the Volga River system. In
this area, the Vyatka has cut into an elevated escarpment of Permian
rocks,
revealing outcrops up to 40 m
high of horizontally banded red, yellow, and brown sediments. These
strata, comprising ancient riverine deposits, have become one of the
best known locations
of
tetrapod fossils.
Along a
relatively straight, south-flowing stretch of the Vyatka River, about
80 km southwest of Kirov, significant Permian
layers with fossil deposits of an age corresponding to the Late
Capitanian stage are exposed near the town
of Kotel`nich. These Middle Permian red beds, composed
of
mudstones, siltstones, marl, sandstones,
and conglomerates, are exposed on the western bank of the Vyatka river
for 24
km between the towns of Kotel`nich and Zemtsy (fig.7). The
Vytaka River is a tributary of the Kama River, which, in turn, is one
of the main branches of the Volga River system. In
this area, the Vyatka has cut into an elevated escarpment of Permian
rocks,
revealing outcrops up to 40 m
high of horizontally banded red, yellow, and brown sediments. These
strata, comprising ancient riverine deposits, have become one of the
best known locations
of
tetrapod fossils.
Fig.7:
Map of the location of the
Kotel`nich Middle Permian deposits, showing geologic
formations (after
Benton et al. 2012)
Fossils from this area were first
recorded by
Krotov
(1894, 1912), who, in mapping the regional geology, found isolated
bones at a locality called
Kotel'nich-1. Little further work was done in the area until twenty
years later, in 1933, when
a young
geologist from Kazan University, S. G. Kashtanov, discovered two
complete
pareiasaur skeletons on the river bank near the village of Vanyushonki,
18 km
south of Kotel'nich. The following year, Kashtanov found two more
pareiasaur
skeletons at an exposure 2 km further upstream , which he sent to the
Paleontological Institute (PIN) in Moscow (Kashtanov 1934).
An expedition led
by A. P. Hartmann-Weinberg, which arrived from Moscow the next year
(1935), found two
incomplete skeletons and two skulls of pareiasaurs near
the
village of Volki. Comparing the initial
findings at
Kotel`nich with those known from Sokolki, Ephremov (1937, 1941)
established two earlier complexes of
dinocephalians, and a later pareiasaurian complex. The latter (equivalent
to the current
Sokolki faunal assemblage; see fig. 2) has
been subdivided into three subassemblages,
with the Kotel'nich fauna now established as the earliest by Ivakhnenko
(1987,
1992),
preceding the Ilyiskoe and Sokolki fauna (Golubev 2000; Benton et al.
2012).
Further
work by the Moscow PIN was delayed until after World War II, when in
1948, a
team led by
B. P. V'yushkov  found
four pareiasaur skeletons near Boroviki village.
The next
year, while searching along a 12 km stretch of the Vyatka River from
Port
Kotel'nich to Boroviki, they discovered thirteen more pareiasaur
skeletons,
six of
these complete (Efremov and V'yushkov 1955).
found
four pareiasaur skeletons near Boroviki village.
The next
year, while searching along a 12 km stretch of the Vyatka River from
Port
Kotel'nich to Boroviki, they discovered thirteen more pareiasaur
skeletons,
six of
these complete (Efremov and V'yushkov 1955).
In the 1990s, work led by D. L. Sumin
from Moscow
PIN recovered a wide variety of tetrapod skeletons, including
dicynodonts,
dromasaurs, therocephalians, and gorgonopsians, as well as pareiasaurs,
of
which 40 skeletons were found the banks of the Vyatka, near the
villages of
Boroviki and Mukha.
Excavations
since
1992 at Port Kotel'nich have
found numerous dicynodont skeletons, which there outnumber those of pareiasaurs.
Along the southern portion of the
banks of the Vyatka both pareisaurs and a unique climbing
reptile named Suminia
have been found. The Vyatka Palaeontological Museum was established in
Kotel'nich in 1994, and has excavated and documented over 390 tetrapod
skeletons.
Plant
fossils found at Kotel`nich include the conifers Phylladoderma
and Geinitzia,
the seed ferms or pteridosperms Tatarina,
Pursongia,
and Permotheca,
and the horsetails Paracalamites
(fig.8)
and Phyllotheca
(Gomankov 1997).
Fig.8: Paracalamitina,
an aquatic plant that grew in Middle and Late Permian riverine
environments
around Kotel`nich.
The
whole sequence at Kotel`nich dates from the middle Tatarian stage, in
the Severodvinian Horizon. This includes two Tetrapod Zones, those
of Deltavjatia
vjatkensis and Chroniosaurus
dongusensis (fig.2).
Based on comparative stratigraphy and magnetostratigraphy,
these correspond to the Late Capitanian stage of the Middle Permian in
South Africa and elsewhere, dated at 263-260 mya (Benton et al. 2012).
The
main reptiles from the
Kotel`nich outcrops include Deltavjatia
(fig.9), large
plant-eating
pareiasaurs whose fossils are often found in groups. Also found are Suminia,
an anomodont with unusually long arms, which may have been one of the
first
arboreal tetrapods. Other members of the Kotel`nich faunal assemblage
are the gorgonopsian Viatkogorgon;
the small insectivore
parareptile Emeroleter (fig.10);
and small carnivorous theriodonts (Benton et al.
2012).
gorgonopsian Viatkogorgon;
the small insectivore
parareptile Emeroleter (fig.10);
and small carnivorous theriodonts (Benton et al.
2012).
The
nearby Port Kotel`nich locality also produced fossils of the
dicynodont Australobarbarus,
as well as two pareiasaurs. Another, somewhat later site in the
immediate area, Sokol’ya Gora, has
yielded the fish-eating
chroniosuchian Chroniosaurus,
the basal biomarsuchian Proburnetia,
and the Pareiasaur Scutosaurus
karpinski, the latter
more
commonly found in Late Permian deposits along the North Dvina river at
Sokolki (Golubev 2000;
Benton et
al. 2012).
Fig.9:
Skeleton of Deltavjayia vjatkensis,
a Middle
Permian pareiasaur from
the
Kotel`nich deposits (after
photo by Khlyupin in
Benton et al. 2012, fig.13).
Equally well represented at Kotel'nich are small, carnivorous
 parareptiles
of the Nycteroleteridae family, of which the type genus,
Nycteroleter,
means "night
thief". Originally
named by
I.A. Efremov in the 1930s for
their large eyes, suggesting night hunting, the
family also includes the genera Emeroleter
("day thief", named
in 1998 by
M.F. Ivakhnenko),
Macroleter
("large thief"), and Bashykroleter.
Their body size is about 20-25 cm, with a relatively large skull 5-7 cm
long.
parareptiles
of the Nycteroleteridae family, of which the type genus,
Nycteroleter,
means "night
thief". Originally
named by
I.A. Efremov in the 1930s for
their large eyes, suggesting night hunting, the
family also includes the genera Emeroleter
("day thief", named
in 1998 by
M.F. Ivakhnenko),
Macroleter
("large thief"), and Bashykroleter.
Their body size is about 20-25 cm, with a relatively large skull 5-7 cm
long.
Fig.10:
Skeleton of Emeroleter (after Khlyupin
2007).
Many examples of Nycteroleter
were found starting with the 1930s excavations by PIN. In recent
excavations sponsored by the Vyatka
Palaeontological Museum, two complete skeletons of the genus
Emeroleter
(fig.10), one adult
and one juvenile, were found together in a fossil
burrow (Khlyupin 2007).
.
The Upper
Tatarian at Mutovino
The
Late Permian exposures at Mutovino, on the Sukhona River (fig.11) have
yielded an unusually rich assemblage of plants, bivalves, insects,
ostracodes, fish, and tetrapods. The locality represents biota
from
the uppermost part of the
Upper Severodvinian Horizon, near its boundary
with the Vyatkian Horizon (fig.2; Golubev 2013).
The Late Tatarian stage of
the East
European Platform is formed mainly of rocks of
lacustrine–alluvial origin The exposure on
the Sukhona River around Mutovino was formed in the
marginal part of the delta of a river that originated on the Baltic
shield (Golubev
2013).
The Sukhona River in this location makes a little northward curve
between the villages of Purtovino and Isady. The northern or left
bank
contains a cliff 50m high and nearly 3 km long, composed of banded
strata of gray and red Permian rocks. In the middle part of this
outcrop, 2.5 km downstream
from the mouth of the Mutovka River, and 1.2 km downstream from the
Mutovino site,
there is a large lens of clay and
siltstone deposits, containing abundant remains, of plants, ostracodes, ,insects, bivalves, fish, and
tetrapods.
Fossil diversity is typically low in this zone, so that a site like
Mutovino with a variety of significant terrestrial and aquatic taxa
provides unusual opportunities for understanding the period (Golubev
2013).
 Geology of the
Mutovino Lens
Geology of the
Mutovino Lens
The locality was formed in the
marginal
part of the delta of a river originating on the Baltic Shield and
flowing into
a large lake located in the northeastern part of the Moscow Syneclise.The Mutovino
lens is located in the middle part
of the riverbank slope. It is clearly visible from a distance, standing
out
against the bluish gray– red banded background of the enclosing
deposits by its
speckled, mostly dark gray coloration. The gray color is a prominent
feature of
this lens, distinguishing it from all other large Permian lenses of the
Sukhona
and Northern Dvina rivers, which usually have yellow– brown
coloration.
At the level of its maximum thickness, the lens
cuts into the underlying deposits to Bed 24; its base is located 9.6 m
higher
than the top of the limestone of Bed 3. The deposits overlying the lens
begin
with Bed 75 (lower beds are indistinguishable; they are either absent
or
facially changed); thus, the maximum thickness of the length is 20 m.
The
position of the edges of the lens could not be determined precisely;
therefore,
the length of the lens was estimated approximately as at least 200 m.
Fig.11:
Map of Late Permian sites
along the Northern Dvina and Sukhona Rivers in western Russia. Most
sites have either of two subassemblanges of the Sokolki Fauna (Solkolki
and Ilinskoe), while another site (Rasha) has the later Vayazniki
Faunal
Assemblage (after Golubev 2000a, fig.6).
The lens
is complex in structure. It is formed
mostly of fine texture matter: clay, siltstone (prevailing), and very
fine to
fine grained sandstone. All rocks are strongly carbonaceous and usually
contain
scattered fish and tetrapod bones and bivalve and ostracode shells. The
upper
part of the lens is formed of siltstones and clays gray, browngray,
brown,
horizontally laminated, containing
fossil plant remains, bivalves, insects, ostracodes, and conchostracans.
Mutovino Fossils
The rocks of the
Mutovino lens contain a diverse assortment of both plant
and
vertebrate fossils. Mutovino is one
of the richest localities of Late
Permian nonmarine organisms in European Russia. Fossils found here
include
abundant plant remains (15 forms), bivalves (six forms), insects (at
least 102
forms), ostracodes (14 forms), conchostracans, fishes (at least seven
forms),
and tetrapods (five forms).
The most
abundant plant fossils include
shoots of the conifers Quadrocladus schweitzeri, in association
with strobili of Dvinostrobus sagittalis. The
subdominant fossils are leaves of the peltasperm pteridosperm Tatarina
conspicua, in association with peltate ovuliphores of Peltaspermopsis,
seeds of Salpingocarpus bicornutus, S. variabilis
Meyen, and sporangia of Permotheca striatifera
and P. vesicasporoides (Golubev 2013).
There are also abundant
fossilized leaves of the
cardiolepids Phylladoderma annulata ,
P. rastorguevii, and P. trichophora In
addition, leaves of the Rhaphidopteris
type have been found. Sporebearing plants are represented by leaves and
megaspores
of the lycapod Lepidophylloides delicata. Other fossil plants
from this locality include leaves with venation of the Taeniopteris
type, assigned to a new fern genus, Fefilopteris.
Abundant fish bones belong to Isadia
sukhonensis A. Minich, (?) I. aristoviensis A. Minich, Isadia
sp., Geryonichthys longus A. Minich, Geryonichthys sp.,
Mutovinia
stella Minich, Strelnia certa A. Minich, Toyemia
tverdochlebovi Minich,
and Actinopterygii ordo indet. (Tatarskie otlozheniya …, 2001).
The tetrapod oryctocoenosis
is represented
exclusively by aquatic and subaquatic forms. The dominant fossils are
the chroniosuchid
anthracosaur Chroniosaurus levis Golubev (63%). Somewhat less
abundant
are the temnospondyl Dvinosaurus primus Amalitzky (26%). The
kotlassiomorphs Microphon sp. and Microphon arcanus
Bulanov occur
less frequently (8%). The predatory therapsid Gorgonopidae gen.
indet.
(2%) and large phytophagous parareptile Pareiasauridae gen. indet. (1%)
are
represented by isolated specimens.
The insect assemblage is
particularly diverse. To
date, the insects described and reported from this locality include
members of
69 families, 81 genera, and 105 species, representing 25 orders.
.
Discoveries
of Late
Permian fossil assemblages on the North Dvina River
In 1892, about the same time as
the fossil beds of Kotel`nich were first investigated, the Russian
geologist Vladimir
P. Amalitzky examined large
concretions found along
the Northern
Dvina River near
Kotlas (fig.11). These concretions were found to contain relativedly
well-preserved fossils of both land vertebrates and plants.
With the
support of the Petrograd Society of Naturalists and the Russian Academy of
Sciences, Amalitzky
and his wife Anna undertook excavations around
Kotlas between 1899 and 1914.
The Late Permian exposures
along the North Dvina and its
tributary the Sukhona River, similar to the Middle
Permian exposures along the Vyatka river to the south, comprise a
series
of thin, superimposed sandstone layers representing ancient river
channel deposits. Within
these loosely-cemented sandstone layers, many
fossils
were found to be contained in the hard, limy concretions
Amalitzky
had initially examined. Since these sometimes
included
whole skeletons as well as bone fragments, it was evident that the
concretions
had formed soon after the death of the animals, while the bones were
still
joined by ligaments. These cacoon-like concretions enabled unusual
levels of preservation. In all, more than 100 tons of
fossil-bearing
concretions
were removed from the North
Dvinian site to
laboratory space
of the Academy of Sciences in St.
Petersburg.
The Dvina River fossils
were found to represent a rich variety of synapsids,
reptiles, and amphibians, as well as freshwater mollusks, and plant
fossils
including Glossoptera. This
Permian conifer is normally found
only in southern latitudes of Gondwanaland, but its occurrence at
various Late Permian sites on the Russian platform, such as those along
the Dvina River, provide a well-known exception.
Amalitzky
died in 1918,
before he
could complete the publication of his reports on the Late Permian
findings
along the North Dvinia River. His
wife Anne
Amalitzky, who had worked closely with him and who drew all the figures
in the
reports, edited his corpus of writings, comprising six mss. on the
paleontology
and three on the geology of the North
Dvinia sites.
In 1921 she
presented the manuscripts to P.A. Pravolslavlev and P.P. Sushkin at the
Russian Academy of
Sciences for final
editing.
They published an initial
portion of
the findings as “The North Dvinia excavations by Professor
Amalitzky,” in two
parts, one on Dvinisaurs and the second on Seymourians. A summary of
the
findings was also published in 1922 in the Bulletin
of the Russian Academy of Sciences (Amalitsky
1922).
Provoslavlev (1927)
thereafter edited and published material from the Amalitzky findings on
gorgonopsians, and Sushkin (1929) on Dvinia
and Dicynodon.
In 1922
the first exhibit
of fossil vertebrates from Kotlas was also
held at the Academy of Sciences in Petrograd.
Exhibited were 10
complete skeletons of pareiasaurs, two
complete skeletons of the gorgonopsian
Inostranzevia,
and several skulls
of
Dicyodon, as well as skeletons of
the primitive Seymouridae
Kotlassia,
the newly discovered amphibian Dvinosaurus,
and
various other vertebrate and plant fossils.
The North
Dvina
excavations revealed numerous remains of Late Permian synapsids which
parallel
both in number and variety those found in the Gondowan deposits of South
Africa, as well
as those known
from Brazil, Australia, Antarctica, India, and China. These
include pareiasaurs, gorgonopsians, anomodonts,
therocephalians, and cynodonts. Each is related to Late
Permian fauna from equivalent
faunal
zones in the South African Beaumont Formation and elsewhere.
 The
pareisaurs of the
Northern Dvinia
region, represented by 13 skeletons, 30 craniums, and many other
isolated
bones,
are somewhat smaller in body
size
than those in the Karoo basin in South Africa, such as Bradysaurus.
The
Russian pareiosaurs are otherwise
notable
for the widely varying sculptured bosses on their skulls, and for
unique
patterns of dermal scales or body armor on their exteriors.
The
pareisaurs of the
Northern Dvinia
region, represented by 13 skeletons, 30 craniums, and many other
isolated
bones,
are somewhat smaller in body
size
than those in the Karoo basin in South Africa, such as Bradysaurus.
The
Russian pareiosaurs are otherwise
notable
for the widely varying sculptured bosses on their skulls, and for
unique
patterns of dermal scales or body armor on their exteriors.
A prominent example is Scutosaurus
karpinski (fig.12),
originally named Pareiosaurus
karpinskii by
Amalitzky
(1922, pp. 334-335). This large animal, 2.45 m in length, had its back
covered with
star- shaped dermal plates, and its belly covered with small conical
bosses. Three related species named
by
Amalitzky were Pareiosaurus
elegans, P.
tubercularum,
and P.
horridus, the latter nearly 3
meters in length, with horn-like
projections
on its cheeks and lower jaws. These three species were later combined
into one, as Scutosaurus
karpinskii
(Ivakhnenko 2008).
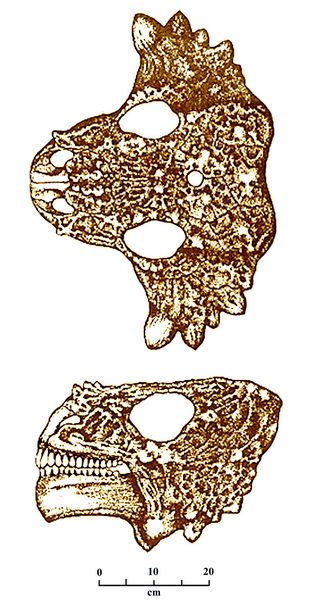
Fig.12:
Skull of Scutosaurus
karpinskii seen from
above and from the left side. The drawing shows intricate patterns of
dermal scales serving as armor.
The theriodonts were
represented by
the large gorgonopsian Inostranzevia
alexandri, of which two complete
skeletons were found, their
craniums 55
and 51 cm. long. These late gorgonopsians had extremely large, serrated
canine
teeth, as well as strong incisors, and conical molars, which were only
in the
upper jaw, the lower jaw lacking any molars (Amalitzky 1922, p.336).
Therocephalians were
represented by
the species Anna Petri
(named for Anna Amalitzky),
with two skulls lacking the lower
jaws.The skull
form was
considered most similar to rhat of Scylacosaurus
in South
Africa (Broom),
but with a
broader and shorter cranium.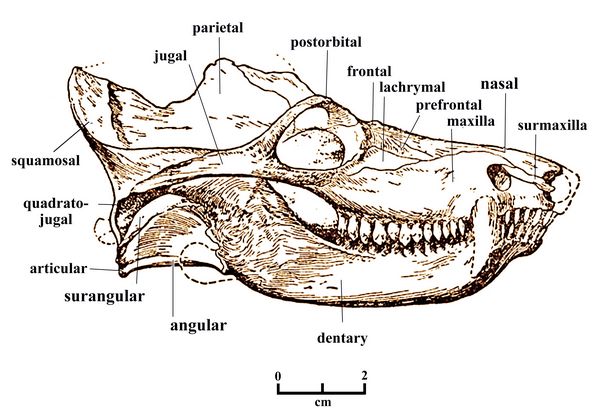
Cynodonts found at Sokolki
include the stem mammal
Dvinia
prima (fig.13), a small
to
medium-sized carnivore with large upper canines, whose
skull
length was 7-10 cm. Dvinia,
initially studied by P.P.
Sushkin,
was more completely defined by Tatarinov (1968).
Dvinia
is considered close to several South African forms from the Late
Permian and
Early Triassic, including Trithelodontia
(Broom), Gomphognathus,
Trirachodon,
and Diademodon.
Fig.13:
Skull of Dvinia
prima, with bones labelled.
Sushkin (1927) also noted
features of
the middle ear region of Permocynodon,
a taxa closely related
to Dvinia,
which Tatarinov (1968, pp.3-6) later redefned as the same species.
These otic
features included a contact between the stapes and the paraoccipital
process,
and the presence of an opening called the foramen stapediale. Tatarinov
placed Dvinia
in the
Procynosuchoid superfamily, and because of its distinct anatomy,
Dvinia
was placed within its own family, Dviniidae. Dvinia
has advanced or
transitional features of both the middle ear and a double set of both
reptilian
and mammalian jaw hinges (Tatarinov 1968, p.33).
The skeletal material of Kotlassia
prima,
a primitive Seymourian
amphibian from the Upper Permian of
the North
Dvina,
was relatively complete. Kotlassia
is
a
terrestrial form, probably similar to Seymouria,
in contrast to the
contemporary Dvinosaurus
(Bystrow 1944). More
recent finds of
related taxa in Khazakstan show they were amphibians and not primitive
amniotes as once thought (Laurin 1996).
Glossary
.
References:
Amalitzky,
V. P.
1922.
"Diagnoses of the new forms of vertebrates and plants from the upper
Permian of North Dvina." Bulletin of
the Russian
Academy of Sciences,
St.
Petersburg
16 (6), pp.329–340.
Benton,
M.J., A.J. Newell, A.Yu Khlyupin, I.S. Shumov, G.D., Price, and A.A.
and Kurkin 2012. "Preservation of exceptional
vertebrate assemblages in Middle Permian fluviolacustrine mudstones of
Kotel'nich, Russia: stratigraphy, sedimentology, and taphonomy."
Palaeogeography,
Palaeoclimatology, Palaeoecology
319-320, pp.58-83
Betekhtina
1966.
“Upper
Palaeozoic non-marine Pelecypoda (bivalves) of Siberia and
Kazakhstan.” Inst.geol.geofiz.,Akad.
Nauk.
Bystrow
A. P. 1944.
"Kotlassia prima
Amalitzky". Geological Society of America Bulletin
55, pp.379-416.
Cisneros,
J.C., F. Abdala, and M.C. Malabarba 2005. "Pareiasaurids from
the
Rio de Rasto Formation, southern Brazil: Biostratigraphic implications
for Permian Faunas of the Parana Basin." Revista Brasileira de
Paleontologia
8(1), pp.13-24.
Efremov,
I.A. and B.P. Vyushkov
1955. “Catalogue of
localities of Permian and
Triassic terrestrial vertebrates in the territories of the
U.S.S.R.” Trudy
Paleontologicheskiy Instituta
46, pp. 1-185.
Golubev,
V.K. 2000a. "The Faunal Assemblages of of Permian Terrestial
Vertebrates from Eastern Europe."
Paleontological Journal 34,
suppl.2, pp. S211-S224.
Ivakhnenko,
M. F. 1981.
"Discosauriscidae
from the Permian of Tadzhikistan." Paleontological
Journal 1981, pp.90-102.
Ivakhnenko,
M. F.
2001.
"Tetrapods from the East European Placket—Late Paleozoic
Natural
Territorial Complex." Proceedings of
the Paleontological Institute of the Russian
Academy of Sciences
283, pp.1–200
Kashtanov,
S.G. 1934. “On the discovery of
Permian reptiles on the River. Vyatka, near the town of
Kotel`nich.”. Priroda
1934 (2), pp. 74-75.
Klembara,
J. 1995. "The
external
gills and ornamentation of skull roof bones of the Lower Permian
tetrapod
Discosauriscus (Kuhn 1933) with remarks to its ontogeny." Paläontologische
Zeitschrift 69, pp. 265-281.
Klembara,
J. & M.
Janiga. 1993.
"Variation in Discosauriscus austriacus (Makowsky 1876) from the Lower
Permian
of the Boskovice furrow (Czech Republic)." Zoological
Journal of the Linnean Society
108. pp. 247-270.
Khimchenko,
L.V., M.N.
Urazaeva, and V.V. Silantiev, 2012. “The microstructure of
aragonite shells of
the non-marine pelecypod genus Concinella
pogorevitsch
Betekhtina 1966.”
Kazan Federal University.
Krotov,
P.I. 1894. “Geological map of European
Russia.” Sheet 89. Trudy
Geologicheskogo Komiteta 13 (2),
pp. 1-228.
Krotov,
P.I. 1912. “The western part of the
Vyatka Province.” Sheet 89. Trudy
Geologicheskogo Komiteta Novaya
Seriya
64, pp. 1-128
Kuznetsov,
V. V. & M.
F. Ivakhnenko.
1981. "Discosauriscids from the Upper Paleozoic in Southern
Kazakhstan." Paleontological Journal 1981:
101-108.
Laurin,
M. 1996. "A
reappraisal of
Utegenia, a Permo-Carboniferous seymouriamorph (Tetrapoda:
Batrachosauria) from
Kazakhstan." Journal
of Vertebrate Paleontology 16
(3), pp.374-383.
Murchison,
R. I.,
1839. The Silurian System.
Murchison,
R. I.,
E. de
Verneuil, and A. von Keyserling, 1845. The
Geology of Russia
in Europe
and the Ural
Mountains.
London,
John Murray.
Newell,
A.J., A. G. Sennikov,
M. J. Benton,
I. I. M Molostovskaya, V. K.
Golubev, A.V. Minikh and
M.
G. 2010.
“Disruption
of playa–lacustrine depositional systems at the
Permo-Triassic
boundary:
evidence from Vyazniki and Gorokhovets on the Russian
Platform,”
Journal
of the Geological Society 167(
4)
,pp. 695-716
Pander,
H.C. 1830. Beiträge
zur
geognosie des russischen reiches
(Contributions to the geology of the Russian Empire).
Pander,
H.C. 1856. Monographie
der Fossilen
Fische des
silurischen Systems der Russisch-Baltischen Gouvernements
(Monograph of fossil
fish from the Silurian stratum of the Baltic regions), St.
Petersburg.
Pander,
H.C. 1857. Ueber
die Placodermen des
devonischen Systems
(On placoderms of the Devonian system). 1857.
Pogorevich,
V.V.
1977. “Some
marine bivalves of the Permian coal-bearing Pechora basin.”
In R.T.
Gratsianova & O.A. Betekhtina (eds.),
New Data About the Fauna and
Stratigraphy of the Middle and Late
Paleozoic of
the USSR, pp. 44-70.
Pravoslavlev,
P. A. (1927). "Gorgonopsidae from the North
Dvinsky
excavations of
V. P. Amalitsky."
Academy
of Sciences
of the USSR,
Leningrad
[St
Petersburg].
Sennikov,
A.G. and V.K
Golubev 2005,`Unique Vyazniki Biotic Complex of the Terminal Permian
from Central
Russia
...` in
The Non-Marine
Permian (ed. Lucas & Zeigler). New Mexico Mus. Nat.Hist
&
Science
Bull.30., pp.302-304..
Silantiev,
V.V.
1998. “New
Data on the Upper Permian bivalve Palaeomutala
in European
Russia.” in Bivalves:
An Eon of Evolution (ed.
Johnston, P.A. & J.W.
Haggart), Univ. of Calvary, Calvary, pp.437-442.
Silantiev,
V.V.
2014.
“Permian Non-marine Bivalve Zonation of the East European
Platform.”
Stratigraphy
and
Geological Correlation,
22(1), pp.1-27.
Silantiev,
V.V. et al. 2013.
The Permian of European Russia.
Kazan Geological Museum, Kazan,
Russia.
Sushkin,
P.P.
1929. “Permocyadon,
a Cynodont reptile from the Upper Permian of Russia.” Proc. X
Internat Congress Zoology, Budapest.
Tatarinov,
L.P.
1968. Morphology
and Systematics of the Northern Dvinia Cynodonts (Reptilia, Synapsida;
Upper
Permian). Yale
University
Postilla 126.
Athena
Review Image Archive™
|
Guide
to Archaeology
on
the Internet |
free trial issue
|
subscribe |
back
issues
Main
index of Athena Review
|
Subject
Index
| Research Pages |
Galleries
aand
Museums |
Ad
rates
|
Current
issue index
Copyright
©
1996-2014
Athena
Publications,
Inc. (All Rights Reserved).