 Triassic
period, before
the rocky plates representing the
continents began to separate.
Triassic
period, before
the rocky plates representing the
continents began to separate.
Middle and Late Permian Therapsids
In
the Permian period, all major land masses were joined in the
mega-continent Pangea,
which formed from the juncture of two
supercontinents, Laurasia
in the north and Gondwanaland in the south. These
had gradually assembled during the preceding Devonian, Mississippian,
and Pennsylvanian
periods. Pangea lasted through the Permian and into
much of
the subequent
Triassic
period, before
the rocky plates representing the
continents began to separate.
A key
indicator of the connection
of land masses in the
Permian
is the distribution of the same species in continental areas now
separated by oceans.
A good example of this is the conifer Glossopteris,
an important Permian forest element preserved in
the fossil
record. Glossopteris
was a
woody, seed-bearing tree, growing as high as 30 meters, and
interpreted to have
thrived in very wet soil
conditions, such as around swamps and along river banks. Fossils
of
Glossopteris
have been found in
all of the southern continents (fig.1), with over 70 species identified
in
Fig.1: Distribution of Glossopteris (green area) in southern Pangea.
The zone of Glossopteris
distribution
(fig.1) across several, now detached,
landmasses led
Austrian geologist Eduard
Suess (1885-1909) to infer that
these areas had once been connected
by a land
bridge, with the continents remaining in the same position.
As
expressed in his major work, The
Face of the Earth
(1885-1909), Suess also believed that
the rise and fall of sea levels were mappable across the earth through
geological time, and that the
periods of ocean transgression and regression could be correlated from
one
continent to another. He
named the interconnected southern land mass
Gondwanaland, after
the district in
Shortly thereafter, the German
meterologist
and polar researcher Alfred
Wegener (1880-1930) more
accurately interpreted that the land
masses themselves had moved together by shifting continental plates.
Wegener's
theory of Continental Drift (1912, 1929), hypothesizing that the
continents were slowly moving around the
Earth, was not widely accepted for several decades. Among its most
vocal opponents was the American paleontologist George Gaylord Simpson
(1943). Wegener's theory began to gain wide acceptance in the
early 1950s, when paleomagnetic samples
taken from
Pangea is the setting for the first appearance in the fossil record of mammal-like reptiles or theriodonts, the ancestors of mammals, during the Middle and Late Permian (268-251 mya). The clearest exposure of this lies in a portion of the southern region of Gondwanaland, centering in South Africa.
.
South Africa: Karoo Basin
The
best continuous fossil record from
the Middle Permian through Early Triassic periods has been found in the
Karoo Basin of South Africa. This sedimentary basin is bounded on the
south by the Cape Fold Mountains, which were uplifted in
the Early to Middle Permian period
by tectonic plates
converging on those of South
Africa (Rubridge 1995). 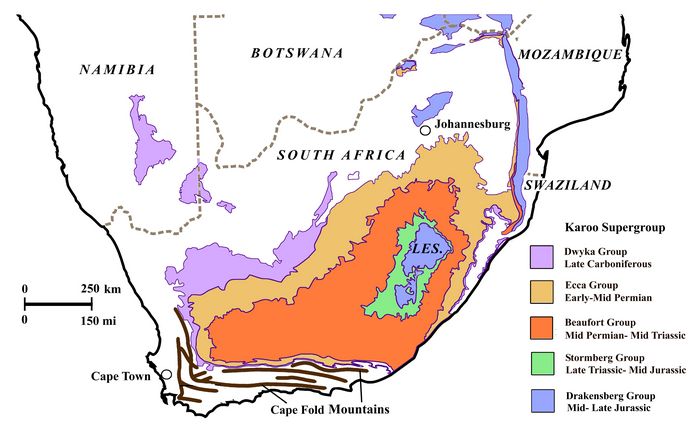
The geological formations in the Karoo Basin include a succession of five groups (Dwyka, Ecca, Beaufort, Stormbert, and Drakensberg), dating from the Late Carboniferous through Jurassic periods (fig.2). The third of these, the Beaufort Group, is the lowest terrestial formation, overlying shallow marine shales of the Ecca Group. Beaufort Group rocks consists of shales and sandstones that represent riverine deposits of an ancient sedimentary basin. Its importance for paleontology stems from its having provided a comprehensive series of often well-preserved fossil zones, including a continuous record of flora and fauna dating from the Middle Permian through Middle Triassic periods (270-220 mya) (Rubridge 1995).
Fig.2: Geological formations in the Karoo Basin of South Africa.
Fossil evidence of Therapsids ("beast faces"), the ancestors of mammals, first appear in the Middle Permian deposits of the Beaufort Group in South Africa. Three suborders of Therapsids will be briefly traced below in their appearance in the Beaufort faunal zones: the Anomodontia ("undefined teeth"), the Dinocephalians ("terrible heads"), and the Theriodonta ("beast teeth"). The theriodonts became one of the two synapsid survivors of the great Permian–Triassic extinction event, the other being the dicynodonts, part of the Anomodontia. Theriodonts split into two groups, Therocephalians ("beast heads") who died out after the Early Triassic; and cynodonts ("dog teeth"), whose carnivorous forms became progressively smaller during the Triassic. By the Late Triassic the first mammals evolved from small, shrew-sized cynodonts called trithelodonts.
The Beaufort group has been subdivided
into eight faunal assemblage zones (A.Z.), each associated with one or
more local geological formations, and each containing a number of
associated taxa (fig.3). 1.
Eodicynodon A.Z., Abrahamskrall Formation; 2.
Tapinocephalus A.Z., Abrahamskraal Formation; 3.
Pristerognathus A.Z., Koonap and Middleton Formations; 4. Tropidostoma
A.Z. Middleton Formation; 5.
Cistecephalus A.Z.,
Middleton
and Balfour Formations; 6.
Dicynodon A.Z., Balfour Formation; 7.
Lystrosaurus A.Z, Balfour and Katberg Formations;
8.
Cynognathus A.Z.,
Burgersdorp
Formation
(Rubridge
1995).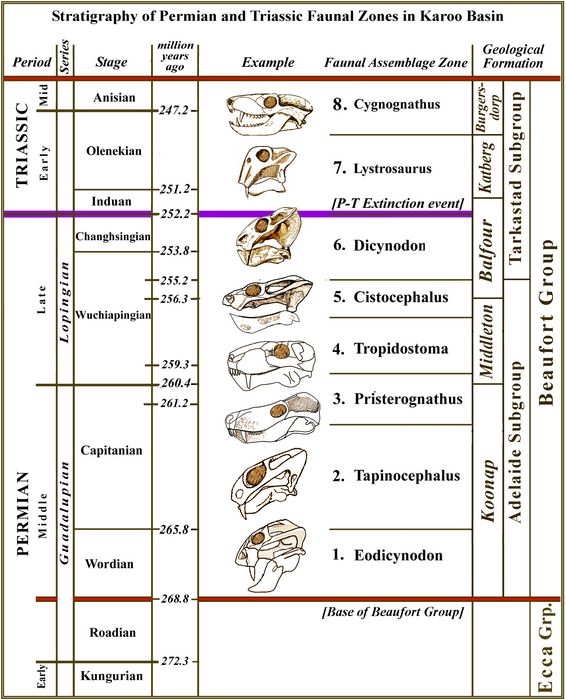
The Abrahamskraal layers,heavily folded to the south, directly overlie marine-deposited shale of the Ecca Group's Waterford Formation. The intersection of the Waterford and Abrahamskraal Formations represents the paleoshoreline at about 270-269 mya (Rubridge 2000; Modesto et al. 2001).
Fig.3: Stratigraphy of the Beaufort Group Faunal Zones. Skulls used as examples: 1. Eodicynodon. 2. Robertia. 3. Pristerognathus. 4. Lycaenops 5. Cistecephalus. 6. Dicynodon. 7. Lystrosaurus. 8. Cynognathus.
The zone
is named for the small
herbivore
Eodicynodon
("dawn
double dog-tooth"), the earliest known genus of
dicynodont, first
found as a skull
at Farm
Zwartskraal near
Dinocephalians:
Anteosauridae
The
huge, semiaquatic
carnivore Anteosaurus
magnificus
("great earlier lizard"), the type species for the
Anteosauridae,
was
named by Watson in 1921.
Up to 5
meters in length, and with a notably long skull
(fig.4), it
had prominent interlocking incisors, large canine teeth, and ten
additional cheek teeth on each side (Boonstra 1954). Anteosaurus probably
lived as a river predator, something like a modern
crocodile. The
giant
Anteosaurus existed
in South Africa at the  same time
that Titanophoneus
and Doliosauriscus
dominated
riverine environments in the Isheevo region in western Russia.
same time
that Titanophoneus
and Doliosauriscus
dominated
riverine environments in the Isheevo region in western Russia.
Fig.4: Skull of Anteosaurus magnificus, from the Mid Permian.
Anteosaurus, defined by a relatively lage sample well-preserved skulls, shows distinctive traits in both its elongated, primitive skull form, and enlarged canine teeth which were related to its carnivorous feeding habits. These traits were passed on to its later descendants such as the Titanosuchidae, many of which, however, changed their dietary habits to become omnivores or herbivores. In terms of their roles as carnivores, the family Anteosauridae were replaced in the Late Permian by large gorgonopsians (described below).
Titanosuchidae
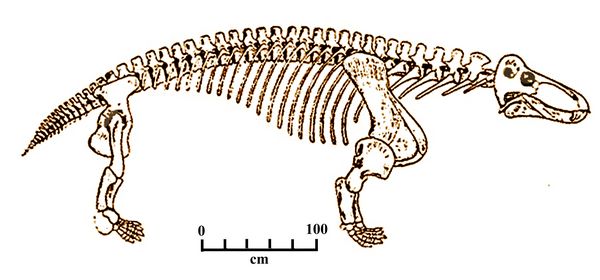
A second genus from the same family, Jonkeria, is thought to have been a partial herbivore, sometimes eating plants (King 1988). Jonkeria ingens (fig.5), defined by Broom (1929), was 4-5 m long, almost as large as Anteosaurus, with long, stout limbs. It had a typically long, dinocephalian snout, with large incisors and canines. A total of six Jonkeria species have been named (Boonstra 1969).
Fig.5: Skeleton of Jonkeria ingens.
Tapinocephalidae
The third
family from
this zone, the
Tapinocephalidae, are
also
known in Russia, and probably had an even wider distribution in the
Middle Permian, before becoming
extinct at the end of the Capitanian stage. Primarily
herbivores, their short, high skulls contrast significantly with the
long, primitive skulls of the prevous two families (Anteosauridae and
Titanosuchidae).
 The
Tapinocephalidae
were among the largest animals of their age, weighting
up to 3,000 - 4,500 pounds, with large, rounded midsections, as is
typical for
plant-eaters. Their teeth have
chisel edges, and (also
in contrast to the previous two families) lack the specialized
canines of carnivores. Their
relatively short, sturdy
forelegs extended outwards, while the longer hind legs were placed
directly under the hips (similar to that of the Late Permian
dicynodonts). Based on comparisons with some modern
herbivores, they could stand bipedally and eat vegetation
directly from lower tree branches.
The
Tapinocephalidae
were among the largest animals of their age, weighting
up to 3,000 - 4,500 pounds, with large, rounded midsections, as is
typical for
plant-eaters. Their teeth have
chisel edges, and (also
in contrast to the previous two families) lack the specialized
canines of carnivores. Their
relatively short, sturdy
forelegs extended outwards, while the longer hind legs were placed
directly under the hips (similar to that of the Late Permian
dicynodonts). Based on comparisons with some modern
herbivores, they could stand bipedally and eat vegetation
directly from lower tree branches.
Tapinocephalus atherstone (fig.6),, the type species for this family, and the only known species of the genus, was a large herbivore up to three meters long, weighing an estimated 3,300-4.400 pounds. Their skulls are characterized by massive frontal bones in the skull roof, and short snouts.
Fig.6: Skull of Tapinocephalus atherstonei, shown in overhead and lateral views (after Smith and Keyser 1995, fig.12a).
Another
large herbivore from the same family
was Moschops
capiensis (Broom 1911). Moschops
ranged from 2.5 - 5.0 m in length, with a large, rounded midsection.
They had
a high, short skull,
with short jaws and chisel-shaped teeth. There are two widely-accepted
species, M.
capensis and M. koupensis,
both found within the same stratigraphic range of the Beaumont
Formation.
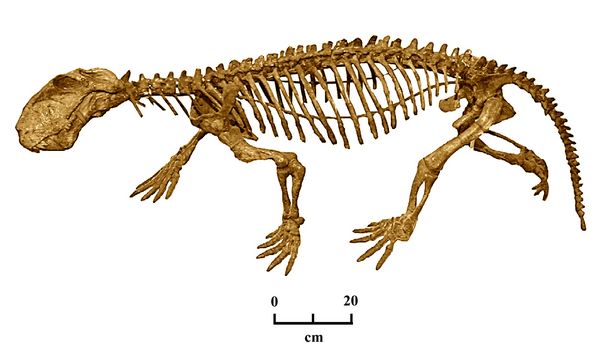
The pareisaurs are represented in this zone by Bradysaurus (figs.7,14), another large herbivore. Bradysaurus had heavily armored scales on its head and neck for protection against gorgonopsians and other predators.
Fig.7: Skeleton of Bradysaurus (Berlin Mus. Nat. Hist.)
The zone is named for Pristerognathus, a medium sized therocephalian carnivore discovered by Broom in 1904, growing up to 1.5 meters in body length. These otter-sized animals had long, narrow skulls with large canines, and probably hunted smaller therapsids and reptiles. Three species include P. baini, P. polyodon, and P. platyrhinus.
At this point, another major change was occurring in the theriodonts,
in the miniaturization of several smaller bones in the rear of the
lower jaw (the reptilian quadrate, articular, and surangular
bones), and their transformation into the tiny ossicle bones
of
the mammalian middle ear (stapes, incus, and malleus). This process,
completed by the Late Triassic, led to the much improved
hearing
of mammals, as compared to reptiles who have only the stapes in the
middle ear to amplify airborne sounds. Improved hearing helped make the
theriodonts the most successful group of synapsids. The two
diagnostic traits of the middle ear with three ossicles, and the
related changes in the lower jaw joint, are the two most commonly used
guides in the fossil record to distinguish early mammals from
non-mammal synapsids as well as reptiles.
4. Tropidostoma
Assemblage
Zone (Late
Permian). This zone, dating from the Late Permian (Tatarian stage) is
named for the early dicynodont, Tropidostoma
("keel mouth"), first
reported by Seeley (1889) and Broom (1915)
from Tafelberg
in the
 the
gorgonopsians show their largest presence, with the greatest number of
their species identified.
the
gorgonopsians show their largest presence, with the greatest number of
their species identified.
6.
Dicynodon Assemblage Zone
(Latest Permian,
257-252 mya). This extensive, ring-shaped zone is
named for Dicynodon
("Two Dog-teeth"), the type species of the dicynodonts, first described
in the mid 19th century. It is a
medium-sized, herbivorous therapsid about 1.2 meters long, with no
teeth except for two prominent, tusk-like canines (fig.8). It probably
cropped vegetation with
a horny beak, much like a tortoise, while the tusks may have been used
for
digging up roots and tubers. The type
species is Dicynodon
lacerticeps, named by Owen
(1845).
Although
over 160 species
of Dicynodon
have since been
named from various Late Permian formations in Russia , China, India,
and elsewhere, these have recently been reclassified. A recent study of
the
genus determined the
only valid members of the genus to be D.
lacerticeps from
South Africa, and D. huenei
from
Fig. 8: Skull of Dicynodon.
7. Lystrosaurus Assemblage Zone (Early Triassic; 252-247 mya). While most of the Permian species described from the previous six faunal zones were eliminated by the massive extinction event at the end of the Permian, some dicynodonts and cynodonts from the Karoo Basin survived, and their fossils are found in Early Triassic layers of the Beaufort Group. The first faunal zone of the Early Triassic in South Africa (fig.6) is named for the small dicynodont Lystrosaurus, a widely dispersed genus who is by far the most commonly found terrestial animal during the Early Triassic. Lystrosaurus will be further discussed in later sections on the Triassic period.8. Cynognathus Assemblage Zone (Early Triassic, 247-242 mya). This zone, located on the inner edge of the Beaufort Group (fig.6), will likewise be discussed in later sections.
.
Late Permian dicynodonts: variation and distributionDicynodonts, well represented in the three Late Permian faunal zones described above for the Karoo Basin of South Africa, and surviving through into the Early Triassic, were a widely distributed group in the southern land areas of Gondwanaland. First appearing in the Middle Permian zones of the Beaufort Group, they became the most successful and abundant land vertebrates of the Late Permian, radiating into a varety of niches in widespread regions as large, medium-sized, and small herbivores, and short-limbed burrowers.
They were first reported in 1845 by
the
geologist Andrew
Geddes Bain from fossils he found during surveys of
As members of the Synapsids, the single temporal opening of dicynodonts is greatly enlarged, supporting very powerful jaw muscles. Their dentition, while somewhat variable, was usually minimal. Many genera, including the type genus Dicynodon, had a complete absence of teeth, apart from their prominent upper canines (fig.8). Some dicynodonts such as Oudenodon had no teeth at all; others, however, such as Pristerodon, had several post canine teeth for chewing, in addition to large canines. One widely distributed genus, Enthiodon, had two rows of 9-11 teeth on the palatine bones of its upper jaw, and matching teeth in its dentary or lower jaw bone, as described by Owen (1879). More consistently, the dentary processes of dicynodonts were made up of a horny, keratin covering on both the upper and lower jaws. This formed a beak-like appearance, resembling that of modern turtles.
Regarding their reproduction, it remains unknown whether dicynodonts laid eggs, as in reptiles, or had developed vivipary, as in mammals. The bony birth canal area between the pubis and ischium was large enough to allow the birth of live young, but could also have been useful in laying large eggs (King 1990).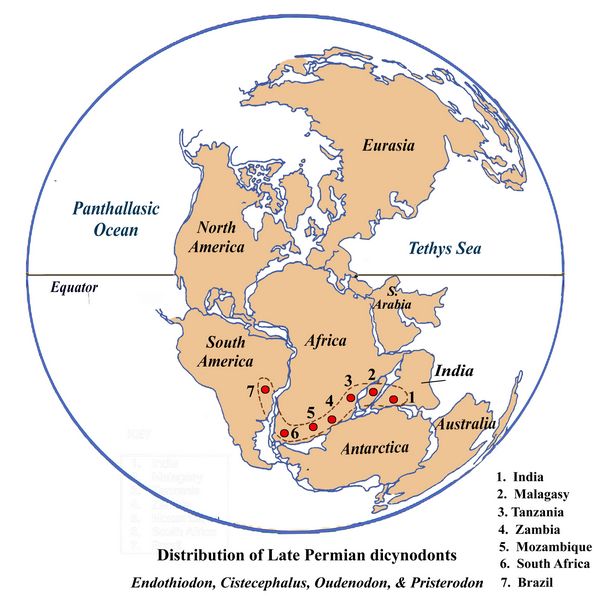 A
distribution
map (fig.9) shows the areas in Gondowanaland where fossils of four
widespread Late Permian dicynodont genera have been found (Ray 1999).
These include Cistecephalus
(already discussed as
the type genus for a faunal zone), Endothiodon, Pristerodon, and Oudenodon. Based on
the overlapping distribution pattern of the widespread conifer Glossopteris, these
animals probably lived in
wooded,
well-watered, riverine settings.
A
distribution
map (fig.9) shows the areas in Gondowanaland where fossils of four
widespread Late Permian dicynodont genera have been found (Ray 1999).
These include Cistecephalus
(already discussed as
the type genus for a faunal zone), Endothiodon, Pristerodon, and Oudenodon. Based on
the overlapping distribution pattern of the widespread conifer Glossopteris, these
animals probably lived in
wooded,
well-watered, riverine settings.
Fig.9: Late
Permian Pangea,
showing distribution of four Late Permian dicynodonts
(after Ray
1999).
Endothiodon
The Late Permian dicynodont Endothiodon (fig.10) was first described by Owen (1879) for the Karoo region, with a skull and mandible, including 11 teeth located in the palatines. A partial skeleton was later recovered by Broom (1915) at Beaufort West in South Africa, from the Hoedemaker member of Middle Teekloof Formation, dating from the Late Permian Tatarian Age. Endothiodon augusticeps is one of the species found in South Africa. Since then, Endothiodon has also been found in Tanzania (Haughton 1932), Zambia (Mazin and King
1991), Mozambique (King 1992), India (Kutty 1970), and Brazil (Boos
et
al. 2013).
also been found in Tanzania (Haughton 1932), Zambia (Mazin and King
1991), Mozambique (King 1992), India (Kutty 1970), and Brazil (Boos
et
al. 2013).
Fig.10: Endothiodon skeleton (after Broom 1915; AMNH) .
The finding of Endothiodon in Brazil is particularly noteworthy, since it is the first dicynodont to be reported for the Permian of South America. A partial skull and associated lower jaw were discovered in the 1970s in a railway cut at Serra do Cadeado, in the Rio do Rasto Formation of Parana State in Brazil (Barbarena and Araujo 1975). The Rio de Rasto Formation is dated at Ufimian to Early Tatarian age, in the Guadelupian Stage of the Middle Permian. The fossils were initially assigned to the genus Endothiodon Owen, which implied a direct correlation of the Brazil specimen with the taxon from the Beaufort Group in South Africa.
Boos et al. (2013) recently reexamined the specimen (FURB PV0226), and have confirmed its original assignment by Barbarena and Araujo to the genus Endothiodon. This is based on several key anatomical features, such as the pineal foramen being located on a boss or small protrusion, a bulbous swellings of the dentaries, a boss situated on the ventral margin of the jugal, an extensive number of dentary teeth, and an upturned pointed beak of the lower jaw (Boos et al. 2013). Further analysis is considered necessary to determine the exact species identification.
Another interesting facet of the findings of Boos
et
al. pertain to biostratigraphical
correlations they propose for the tetrapod faunas of the Rio do Rasto
Formation. These show similarities with stratigraphic ranges of the
Middle
and Late Permian of both South Africa and Eastern Europe.
As Boos et al. (2013) state, the presence of Endothiodon
shows that part of the
Rio do
Rasto Formation in Brazil can now be correlated with deposits in India,
Malawi, Mozambique, South Africa, Tanzania, Zambia, and Zimbabwe.
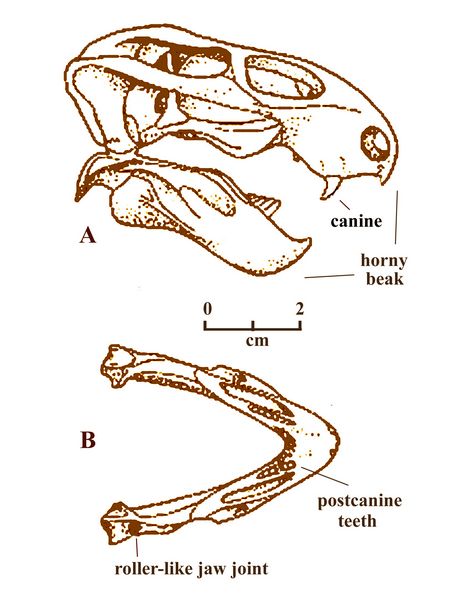 skull
4-6 cm long, with a short snout (fig.11). Unlike
most other dicynodonts, Pristerodon
retained six pointed post-canine teeth on either side of its upper and
lower jaws (maxilla and dentary). The rest
of the jaws were made of sharp bone, probably covered with keratin or a
horny surface (Benton 2005). Pristerodon
lived on
vegetation which it snipped off with its horny beak, then passed back
to its cheek area for grinding with its molars before
swallowing.
skull
4-6 cm long, with a short snout (fig.11). Unlike
most other dicynodonts, Pristerodon
retained six pointed post-canine teeth on either side of its upper and
lower jaws (maxilla and dentary). The rest
of the jaws were made of sharp bone, probably covered with keratin or a
horny surface (Benton 2005). Pristerodon
lived on
vegetation which it snipped off with its horny beak, then passed back
to its cheek area for grinding with its molars before
swallowing. Fig.11: Pristerodon skull (A) and lower jaw (B), showing teeth (after Benton 2005)
Oudenodon
Oudenodon
is the type genus of the
family Oudenodontidae, which
also includes Cteniosaurus,
Tropidostoma,
and Rhachiocephalus.
The
Oudenodontidae family have a deep, thin
walled cleft in the lower jaw for anchoring the horny platform.  They
had a very
precise cutting and crushing surface between the beaks, where the rim
of the
maxilla was pointed downward to fit with the lower jaw symphesis. Oudenodon
(fig.12) had
no teeth at all. The skull was directed forwards, therefore, adapted to
feeding
at levels approximately 20-100 cm above the ground (King 1990). This
shows a
new level of diversification, away from substrate-feeding.
They
had a very
precise cutting and crushing surface between the beaks, where the rim
of the
maxilla was pointed downward to fit with the lower jaw symphesis. Oudenodon
(fig.12) had
no teeth at all. The skull was directed forwards, therefore, adapted to
feeding
at levels approximately 20-100 cm above the ground (King 1990). This
shows a
new level of diversification, away from substrate-feeding.
Fig.12: Oudenodon skull,
showing beak-like
jaws (Berlin Museum of Natural History)
Oudenodon was
common throughout
southern Africa during the Late Permian, and has also been found in
India (Ray
1999; Bandyopadhyay
1999),
Madagascar,
and Zambia. Several
known species of Oudenodon
include Oudenodon bainii,
the type species, and O. grandis from
South Africa, and O. luangwensis
from Zambia. Another species, O.
sakamenensis, is the only
therapsid yet known
from Madagascar (Mazin and King 1991).
Gorgonopsians: the main
Late
Permian predators
Gorgonopsians (Gorgon and -ops-,
"eye" or "face" = "Gorgon
faces"), are the most primitive of the
three groups of theriodonts found in the Karoo Basin, the other
two being therocephalians and cynodonts. During the Late Permian,
the gorgonopsians were a highly successful group, including a total of
25 genera and 41 species, from South Africa,
Tanzania, Zambia, and Malawi, as well as from Russia (Sigogneau-Russell
1989; Ivokonenko 2005; Gebauer 2007). 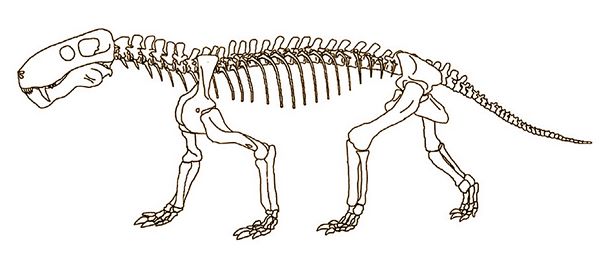
In
Fig.13: Skeleton of Sauroctonus, a Late Permian gorgonopsian found in Russia and Africa (Gebauer 2007)
During the Late
Permian, gorgonopsians became much more common, with the genera Gorgonops,
Eoarctops,
Lycaenops,
Sauractonus
(figs.13,16) Arctognathus,
Njalila,
Clelandina, and
Rubidgea
being found in the
Tropidostoma and
Cistecephalus Zones dated at 260-255 mya (Rubidge 1999; Smith and
Keyser 1995).
In the Dicynodont Zone (255-252 mya) occur the latest forms of
Gorgonopsians, including
Cynosaurus, Prorubidgea,
and Rubidgea, the latter with
extremely enlarged canines. Gorgonopsians
then became extinct as part of the mass extinction at the end of the
Permian (251 mya), with the
other two
theriodont groups continuing into the Triassic (Rubidge et al.
1995; Gebauer 2007).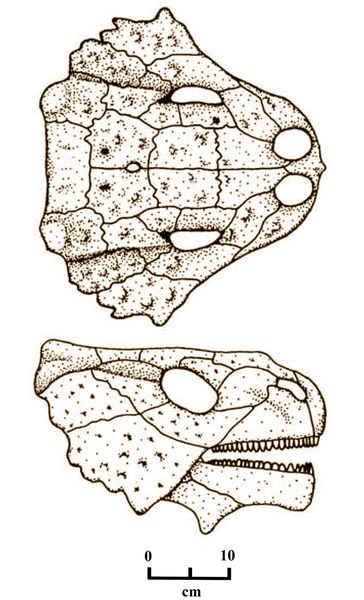
Fig. 14: Skull of Bradysaurus seen from above and from the side, showing the rough, scaly armor used as a defense against predators (after Smith and Keyser 1995).
Gorgonopsians were among the
largest
carnivores of the Late Permian. Most of the known forms, such as the
type genus Gorgonops,
were 1.5 - 2 meters in
length, about the size of lions and other large felids. The largest
known genus was Inostrancevia,
found in Late Permian deposits in the Northern Dvina
Valley in Russia, in the Sokolki assemblage (Efremov
1937; Ivakhnenko
1990). Inostrancevia
was
over 3 meters long, about the size of the largest modern bears, with a
skull length
of 45 cm, and saber-like canine teeth
up to 12
cm long.
The mammal-like traits of gorgonopsians include differentiated or heterodont teeth, with incisors, canines, and cheek teeth or molars; a large, fully developed temporal fenestra for strong jaw muscle attachment; and vertically-aligned rear legs for faster movement. Gorgonopsids also had a vaulted palate that may have facilitated breathing while holding prey in their jaws; and early stages of the hearing ossicles that later developed in the mammalian middle ear.
The type
species, Gorgonops
torvus, was first identified on
the basis of an incomplete and
flattened
skull found in South Africa at Mildenhalls,
 The
type genus
Gorgonops, like most members of the family gorgonopsidae, has a
body length of 1.2-2
meters and a
skull length of 22-35 cm. Its formidable array of teeth included the
large canines, which had serrated edges, and five incisors. Besides
G.
torvus, among the earlier
species to be described were the larger Gorgonops
whaitsi (Broom, 1912), found in the
Tropidostoma and Cistecephalus Zones in the
Beaufort West area. Sigogneau-Russell
(1989)
considered this a more primitive form
than G.
torvus. A third and still
larger
species, which
may be descended from G.
whaitsi, is
Gorgonops
longifrons (Haughton 1915),
known from an incomplete and
flattened skull
about 35 cm long, with larger eye orbits and a longer snout.
The
type genus
Gorgonops, like most members of the family gorgonopsidae, has a
body length of 1.2-2
meters and a
skull length of 22-35 cm. Its formidable array of teeth included the
large canines, which had serrated edges, and five incisors. Besides
G.
torvus, among the earlier
species to be described were the larger Gorgonops
whaitsi (Broom, 1912), found in the
Tropidostoma and Cistecephalus Zones in the
Beaufort West area. Sigogneau-Russell
(1989)
considered this a more primitive form
than G.
torvus. A third and still
larger
species, which
may be descended from G.
whaitsi, is
Gorgonops
longifrons (Haughton 1915),
known from an incomplete and
flattened skull
about 35 cm long, with larger eye orbits and a longer snout.
Fig.15: Skeleton of Lycaenops, a Late Permian gorgonopsian found in the Beaufort Group (photo: after Gebauer 2007)
Lycaenops ("Wolf-face"), found in the Late Permian Endothiodon Zone, was another medium-sized gorgonopsian, with more enlarged canines. The first named species of this genus was Lycaenops ornatus (fig.15), whose type specimen was a nearly complete skeleton found by Broom in 1920 on a weathering shale slope near a railway line, two miles south of the railway station at Biejespoort, South Africa. Stratigraphically, it was at the very top of the Endothiodon zone, within the Teekloof Formation of the Karoo Basin. Some carpal bones of the hands and feet had weathered away along with most of the tail and cervical region, but otherwise the skeleton was intact, a rarity for Gorgonopsia fossils (Broom 1930, pp.349-50; Colbert 1948).
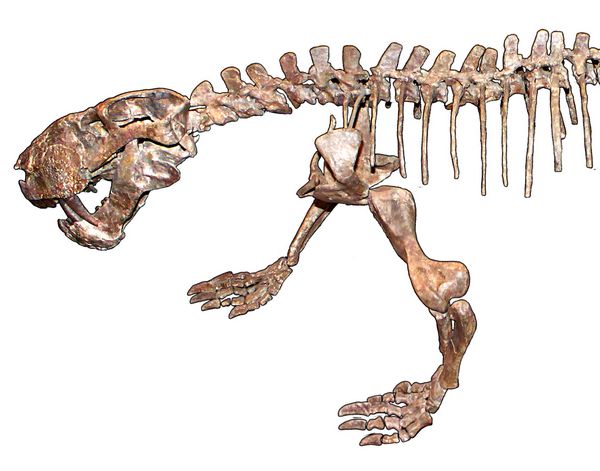 Regarding
the
undoubted identity of gorgonopsians as Late
Permian "saber-toothed" carnivores, Colbert
observed that the condition of hypertrophy of the canines, accompanied
by a deepening of the anterior part of the lower jaw, is well
represented in Lycaenops ornatus,
but is seen at its
most advanced form in the late gorgonopsians Inostrancecia
and Rubidgea
(Colbert 1948, pp.373-375). More recently, Gebauer
(2007) has made a detailed
comparison between the
gorgonopsian Sauroctonus (figs.13,16),
first described from the Northern Dvina region in Russia (Tatarinov
1974), and
the saber-toothed cat Smilodon
fatalis (fig.17), which had
similar body lengths of about 2 meters.
Regarding
the
undoubted identity of gorgonopsians as Late
Permian "saber-toothed" carnivores, Colbert
observed that the condition of hypertrophy of the canines, accompanied
by a deepening of the anterior part of the lower jaw, is well
represented in Lycaenops ornatus,
but is seen at its
most advanced form in the late gorgonopsians Inostrancecia
and Rubidgea
(Colbert 1948, pp.373-375). More recently, Gebauer
(2007) has made a detailed
comparison between the
gorgonopsian Sauroctonus (figs.13,16),
first described from the Northern Dvina region in Russia (Tatarinov
1974), and
the saber-toothed cat Smilodon
fatalis (fig.17), which had
similar body lengths of about 2 meters.
Fig.16: Skull
and anterior portion of the
skeleton of Sauroctonus parringtoni,
from the Upper Permian in the
In spite of the fact that they are separated by at least 230 million years, and that Sauroctonus still retained some reptilian features in its lower jaw, skull, and skeleton, while Smilodon was an advanced mammal, there are some intriguingly close functional parallels between these two extinct carnivores, both in teeth and jaw forms, and the musculature of jaw movements used in catching and eating prey.

The lower jaws of gorgonopsians, for example, had a thicker front portion than the rear portion, protecting the enlarged canine teeth; a similar function was performed by bone flanges of saber-toothed cats. After a detailed analysis, Gebauer concludes that "it can be well imagined that Sauroctonus in all probability occupied the same ecological niche [as] its mammalian relative 230 million years later" (Gebauer 2007).
Fig.17: Skeleton of the saber-toothed cat Smilodon fatalis (after Gebaur 2007).
Gorgonopsians in Tanzania, Malawi, and Zambia
Haughton (1927) later described two gorgonopsian genera found in Malawi, Gorgonops and Aelurognathus (fig.18), the latter genus previously named by Owen (1881). Boonstra (1934), based on analysis of post-cranial anatomy, described two species of the genus Aleruognathus, and one of the genus Arctognathus.
Boonstra
(1953) later described gorgonopsians
from Tanzania, as did Parrington
(1955, 1972), including those
found by the German engineer Nowack in the 1930s. Identified taxa
from Tanzania
now
include
Aloposaurus,
Leontocephalus,
Dinogorgan,
Sauroctonius,
and possibly Arctognathus (Gebauer
2007). Including
the
Tanzanian
materials,  Kemp (1982, 2005) has made
detailed
comparisons
of gorgonopsian
skull and skeletal anatomy with other synapsids. More
recently, Gebauer
(2007) reanalyzed one of the
best preserved of the Tanzanian specimens, a nearly complete skeleton
originally described as Aelurognathus,
now identified as a variant of
the
Russian genus Sauroctonius
(Tatarinov 1974).
Kemp (1982, 2005) has made
detailed
comparisons
of gorgonopsian
skull and skeletal anatomy with other synapsids. More
recently, Gebauer
(2007) reanalyzed one of the
best preserved of the Tanzanian specimens, a nearly complete skeleton
originally described as Aelurognathus,
now identified as a variant of
the
Russian genus Sauroctonius
(Tatarinov 1974).
Fig.18: Skull of Aelurognathus, a Late Permian gorgonopsian found iin Tanzania, Malwai, and Zambia, as well as South Africa.
From Zambia, specimens have been identified as Aelurognathus by Sigogeneu (1970). In general for the gorgonopsians, many problems in redundant classifications (such as addressed by Colbert in 1948) were sorted out by Sigogneau-Russell (1989), although some difficulties remain, given the number of gorgonopsian taxa and the wide geographical range of their presence.
The most primitive known gorgonopsians
are
pelycosaur-like forms in Late Permian Russia, including Biarmosuchus
and Eotitanosuchus
from
Ezhevo.
They are contemporary with more
advanced forms from South Africa, and thus can provide no simple
answers to the origins of gorgonopsians. Other, more derived Russian
forms include Sauroctonius, Viatkogorgon,
and Suchogorgon
(Tatarinov 1974, 1999,
2000; Ivakhnenko
1990). These and other Russian
taxa are discussed in the following section.
.
.Anderson, J.M., and A.R. Cruikshank 1978. "The Biostratigraphy of the Permian and Triassic. Part 5: A review of the classification and distribution of Permo-Triassic tetrapods." Palaeontologia Africana 21, pp. 15-44.
Bain,
A.G. 1845. "On
the
discovery of fossil remains of bidental and other reptiles in
Barberena, M.C., D.C. Araujo, and E.L. Lavina 1985. "The evidence for close paleofaunistic affinity between South America and Africa as indicated by Late Permian and Early Triassic tetrapods." In Ulbick, H.Y. and A. Rocha-Campos (eds.), Gondwana Proceedings, 7th International Gondwana Symposium, San Pablo, pp. 454-467.
Barry, T.H. 1974. "A new dicynodont ancestor from the Upper Ecca." Annals of the South African Museum 64, pp.117-136.
Boonstra, L.D. 1938. "A report of some Karoo reptiles from the Luangwa Valley, Northern Rhodesia" Quaternary Journal of the Geological Society of London 94, pp. 371-384.
Boonstra,
L.D. 1953
"A report on a
collection of fossil reptilian bones from
Boonstra,
L.D. 1954."The cranial structure
of the Titanosuchian Anteosaurus."
Ann. S.
Afr. Mus.,
Boonstra,
L.D. 1969."The fauna of
the Tapinocephalus Zone (Beaufort Beds of the
Boos, A. D. S.; Schultz, C. L.; Vega, C. S.; Aumond, J. S. J. 2013. "On the presence of the Late Permian dicynodont Endothiodon in Brazil". In Angielczyk, Kenneth.
Broom, R. 1912. "On some new fossil reptiles from the Permian and Triassic beds ofBroom, R.
1913a.
"On a nearly perfect skull of a new
species of the Gorgonopsia." Ann.
S. Afr. Mus., 12, pp. 8-10.
Broom, R 1913b.
"On the Gorgonopsia, a sub-order of the
mammal-like reptiles." Proc.
zool. Soc. London, 1, p.. 225
Broom,
R. 1925. "On some carnivorous
therapsids." Rec. Albany
Mus., 3. pp. 309-326.
Broom, R.
1930. "On the
structure of
the mammal-like reptiles of the sub-order Gorgonopsia" Phil. Trans. Roy. Soc.
Broom, R.
1940. "On some
new genera
and species of fossil reptiles from the
Colbert,
E.H. 1948.
"The mammal-like
reptile Lycaenops."
Bull.
Am. Mus. Nat. Hist., 89, pp.
357-404.
Efremov,
I.A. 1937. "On the
stratigraphic
division of the
continental Permian and Triassic USSR on the basis of the terrestrial
vertebrate fauna."
–
Dokl. Akad. Nauk SSSR, Nov. Ser.,
16(2), pp.125-132.
Gebauer, E.V.I. 2007 . "Phylogeny and evolution of the Gorgonopsia with a special reference to the skull and skeleton of GPIT/RE/7113 ('Aelurognathus?' parringtoni)" Ph.D. thesis, Tübingen: Eberhard-Karls Universität Tübingen. pp. 1–316.
Gregory, W.T. 1926. "The skeleton of Moschops capensis Broom, a dinocephalian reptile from the Permian of South Africa." Bull. Amer. Mus. Nat. Hist. 56: 179-251
Haughton, S.H. 1915. "Investigations in South African fossil reptiles and Amphibia, 7. On some new gorgonopsians." Ann. S. Afr. Mus.,12: 82-90.
Haughton, S. H. 1917. "Investigations in South African fossil reptiles and Amphibia. Part 10. Descriptive catalogue of the Dicynodontia." Annals of the South African Museum 12, pp.127-174.
Haughton,
S.H. 1927.
"On
Ivakhnenko,
M.F. 1990.
"The
late Palaeozoic faunal assemblage of tetrapods from deposits of the
basin of
the
Ivakhnenko,
M.F. 2005.
"Comparative survey of Lower Permian
tetrapod faunas of
Jacobs, L. L., Winkler, D. A., Newman, K. D., Gomani, E. M. & Deino, A., 2005, "Therapsids from the Permian Chiweta Beds and the age of the Karoo Supergroup in Malawi." Palaeontologia Electronica. Vol. 8, #1, pp. 28A: 21-23.
Kammerer, C.F. and K.D. Angielczyk 2009. "A proposed higher taxonomy of anomodont therapsids". Zootaxa 20, pp. 1–24
Kemp,
T.S. 1982.
Mammal-like
reptiles and the
origin of mammals. Academic
Press.,
Kemp,
T.S. 2005. The Origin and
Evolution
of
Mammals. Oxford University
Press, pp.331.
Keyser, A. W. 1975 . "A re-evaluation of the cranial morphology and systematics of some tuskless Anomodontia." Memoir of the Geological Society of South Africa 67, pp.1-110.
King, G.M. 1988. "Anomodontia. Encyclopedia of Paleoherpetology, Part 17c, Gutsav Fischer Verlag, Stuttgart & New York.
King,
G.M. 1990. The Dicynodonts: A
Study in Palaeobiology.
Chapman & Hall.
King, G.M. 1992. "The paleobiogeography of Permian anomodonts." Terra Nova 4, pp.633-640.
Kutty, T.S. 1972. "Permian reptile fauna from India." Nature 237, pp.462-463.Ochev, V.G., 2001. Zonal stratigraphic correlations of the Upper Permian from the Cis-Urals and South Africa according to tetrapods. 6th European Workshop on Vertebrate Palaeontology - Florence and Montevarchi.
Olson, E.
C. 1937.
"The
cranial morphology of a new gorgonopsian." J. Geol.,
45, pp.511-527.
Owen, R.
1876a. "Evidences of
theriodonts in Permian deposits elsewhere than in
Owen, R.
1876b
Descriptive
and Illustrated Catalogue of the Fossil Reptilia of
Owen, R. 1881. "On the order Theriodontia with a description of a new genus and species (Aelurognathus fel. ow.)." Quat. Jour. Geol. Soc. London, 37, pp. 261-265.
Parrington, F.R. 1955. "On the cranial anatomy of some gorgonopsids and the synapsid middle ear." Proc. Zool. Soc.Seeley,
H.G. 1895.
"Researches on
the structure, organization and classification of the fossil Reptilia.
IX,
section 1. On the Therosuchia." Phil.
Trans. Roy. Soc. London, B,
185, pp.
987-1018.
Smith,
R.H.M. and Keyser,
A.W.
1995. Biostratigraphy of the Tropidostoma Assemblage Zone. Geological
Survey of
Suess,
Eduard
1885-1909. Das Antlitz der
Erde [The Face of the Earth].
F. Tempsky,
Tatarinov,
Tatarinov,
I. P.
1999. "New
Theriodonts (Reptilia) from the late Permian Fauna of the town of
Tatarinov,
Watson,
D.M.S. 1914.
"Notes on
some carnivorous reptiles." Proc.
Zool. Soc. London, 4,
pp.1021-1038.
Watson,
D.M.S. 1921. "The
bases
of classification of the Theriodonta" Proc. Zool. Soc. London,
1, pp.
34-98.
Watson, D.M.S. 1948. "Dicynodon and its allies." Proc. Zool. Soc. London, 118, pp.823-877.
Wegener,
Alfred 1912. "Die
Entstehung der
Kontinente"
[The formation of continents] Geologische
Rundschau, Zeitschrift für allgemeine
Geologie , dritter Band, pp.
276-292
Wegener,
Alfred 1929 . Die
Entstehung der Kontinente und Ozeane [The Origin of Continents
and Oceans]
(4th ed.). Braunschweig: Friedrich Vieweg & Sohn Akt.
Wegener,
Alfred 1966. The Origin
of Continents and Oceans.
Willem A.
J. M. et
al. 1928. Theory of Continental
Drift: a Symposium
of the Origin and Movements of Land-masses of both Inter-Continental
and Intra-Continental,
as proposed by Alfred Wegener.
Athena Review Image Archive™ | Guide to Archaeology on the Internet | free trial issue | subscribe | back issues
Main
index of Athena Review
|
Subject
Index
| Research Pages |
Galleries
aand
Museums |
Copyright © 1996-2014 Athena Publications, Inc. (All Rights Reserved).